mRNA 的修饰方式有哪些?它们分别作用于哪些位置?
mRNA 修饰是基因表达调控的关键环节,通过改变结构影响其转运、翻译与降解。这些修饰分布于不同位置,5’ 端有帽子结构,3’ 端存在多聚腺苷酸化,内部还有甲基化等多种方式,共同调控 mRNA 生命周期。
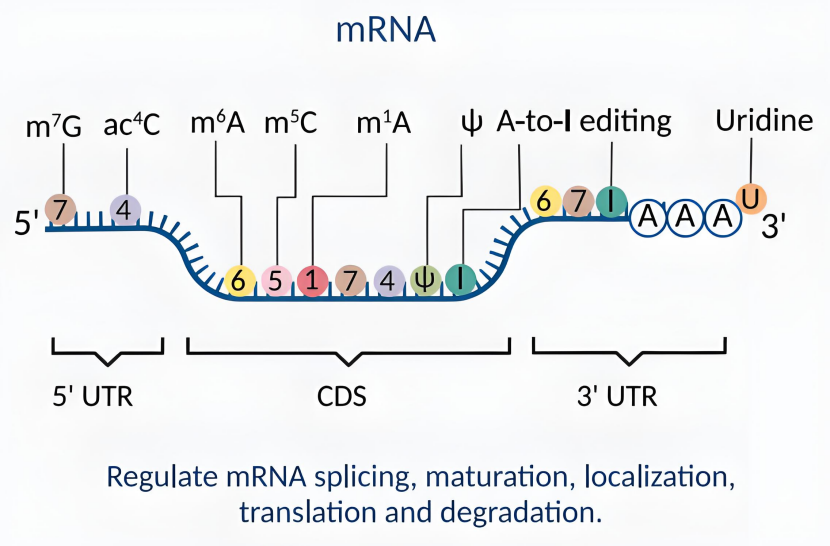
5’ 端修饰:帽子结构(Cap)
mRNA 的 5’ 端修饰是转录起始后不久发生的关键事件,由一系列酶催化形成,核心功能是保护 mRNA 免受 5’→3’ 核酸酶降解,并促进翻译起始。
01
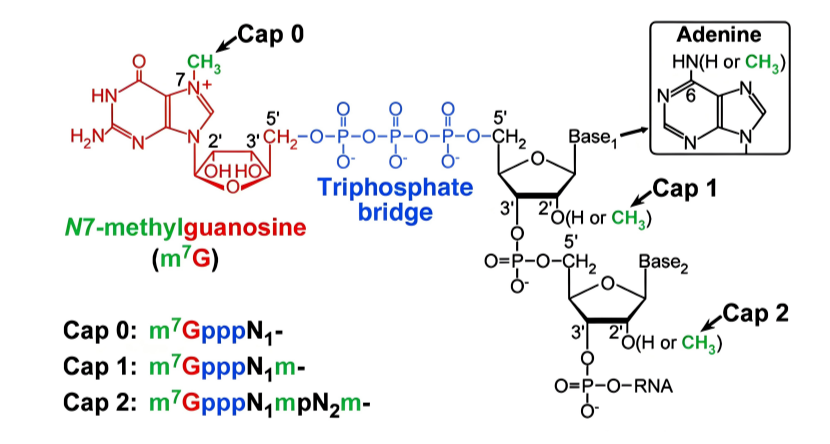
帽子结构的核心组成
5’ 端第一个核苷酸(通常是鸟苷酸,G)通过 5’-5’ 三磷酸键与 mRNA 的第一个核苷酸连接(不同于常规的 3’-5’ 磷酸二酯键),形成 “GpppN” 结构(N 为 mRNA 的第一个编码核苷酸)。
鸟苷酸的第 7 位氮原子被甲基化(-CH₃),形成7 - 甲基鸟苷帽(m⁷G),这是所有帽子结构的共同特征。
02
帽子的类型(基于甲基化程度)
**Cap 0:**仅 7 - 甲基鸟苷(m⁷G),第一个核苷酸的 2’-OH 未甲基化(常见于低等真核生物,如酵母)。
**Cap 1:**m⁷G + 第一个核苷酸的 2’-OH 甲基化(m⁷GpppNm,“Nm” 表示 2’-O - 甲基核苷酸),是高等真核生物(如哺乳动物)的主要帽子类型。
**Cap 2:**m⁷G + 第一个和第二个核苷酸的 2’-OH 均甲基化(m⁷GpppNmNm),存在于部分高等真核生物 mRNA 中,功能尚不完全明确。
03
生物学功能
与帽结合蛋白(eIF4E)结合,介导核糖体小亚基组装,启动翻译。
阻止 5’→3’ 外切酶降解 mRNA,延长其半衰期。
促进 mRNA 从细胞核转运到细胞质。
3’ 端修饰:多聚腺苷酸化(Polyadenylation)
mRNA 的 3’ 端修饰是转录终止后通过 “切割 - 加尾” 过程形成的,核心是添加一段多聚腺苷酸(poly (A))尾,长度通常为 50-250 个腺苷酸(A)。
01
形成机制
转录生成的前体 mRNA(pre-mRNA)在 3’ 端的特定位点(由 poly (A) 信号序列 “AAUAAA” 和下游 GU 富集序列界定)被内切酶切割。
由多聚腺苷酸聚合酶(PAP)催化,在切割后的 3’ 端添加连续的 A 残基,形成 poly (A) 尾。
02
生物学功能
与 poly (A) 结合蛋白(PABP)结合,协同 5’ 端帽子结构稳定 mRNA(形成 “闭环” 结构),抑制 3’→5’ 外切酶降解。
增强翻译效率(PABP 与翻译起始因子相互作用)。
参与 mRNA 的核输出和细胞定位。
内部碱基修饰:甲基化
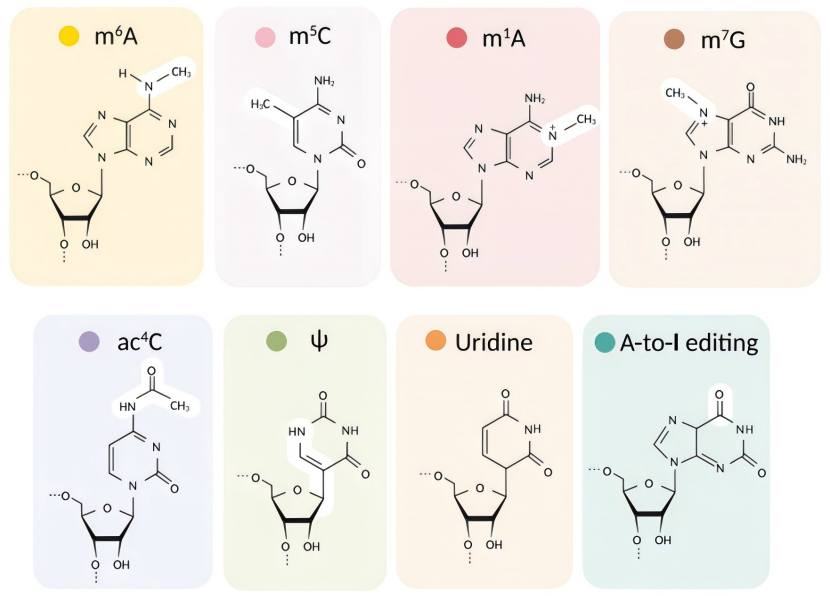
mRNA 的内部(编码区或非翻译区,UTR)碱基可发生甲基化修饰,其中**N⁶- 甲基腺苷(m⁶A)**是最常见且研究最深入的类型。
01
N⁶- 甲基腺苷(m⁶A)
**结构:**腺苷(A)的第 6 位氮原子被甲基化(-CH₃)。
**分布:**主要集中在 mRNA 的编码区(CDS)和 3’UTR,尤其在 “RRACH” 序列(R=A/G,H=A/C/U)中富集。
调控因子:
-
**甲基转移酶(Writers):**如 METTL3/METTL14 复合物,负责催化 m⁶A 形成;
-
**去甲基化酶(Erasers):**如 FTO、ALKBH5,可去除甲基,使修饰可逆;
-
**结合蛋白(Readers):**如 YTHDF 家族,通过识别 m⁶A 调控 mRNA 的翻译或降解。
**功能:**影响 mRNA 剪接(如促进某些外显子 inclusion);调控翻译效率(如 YTHDF1 结合 m⁶A 后增强翻译);促进 mRNA 降解(如 YTHDF2 介导 mRNA 进入降解复合体)。
02
其他甲基化修饰
**N¹- 甲基腺苷(m¹A):**腺苷第 1 位氮原子甲基化,主要位于 mRNA 的 5’UTR 和起始密码子附近,可能增强翻译起始。
**5 - 甲基胞嘧啶(m⁵C):**胞嘧啶(C)第 5 位碳原子甲基化,分布于编码区和 3’UTR,功能与 mRNA 稳定性和翻译相关(如在肿瘤细胞中可能上调致癌基因翻译)。
**7 - 甲基鸟苷(m⁷G):**除 5’ 端帽子外,内部 G 也可能被甲基化,参与 mRNA 的核输出和翻译调控。
假尿苷化(Pseudouridylation,Ψ)
假尿苷(Ψ)是尿苷(U)的异构体,由尿苷的糖苷键重排形成(尿嘧啶的 C5 与核糖的 C1 连接,而非常规的 N1)。
01
形成机制
由假尿苷合酶(PUS 家族)催化,无需模板,直接将 mRNA 中的 U 转化为 Ψ。
02
功能
增强 mRNA 的稳定性(Ψ 比 U 更难被降解);
改变 RNA 的二级结构(增加碱基配对能力),影响翻译效率;
在应激条件下(如热休克),mRNA 的假尿苷化可能上调应激相关蛋白的表达。
碱基编辑(Base Editing)
mRNA 的碱基编辑是指特定核苷酸被化学修饰后转变为另一种碱基,直接改变密码子信息,影响翻译产物。
01
腺苷脱氨基编辑(A-to-I)
由腺苷脱氨酶(ADAR 家族)催化,将腺苷(A)脱氨基为肌苷(I)。
由于肌苷在翻译时被识别为鸟苷(G),可导致密码子改变(如 AUA→IUA→GUA,对应氨基酸从异亮氨酸变为缬氨酸)。
**例子:**谷氨酸受体(GluR)mRNA 的 A-to-I 编辑可改变通道的钙离子通透性,影响神经信号传导。
02
胞嘧啶脱氨基编辑(C-to-U)
由胞嘧啶脱氨酶(APOBEC 家族)催化,将胞嘧啶(C)脱氨基为尿嘧啶(U)。
**例子:**载脂蛋白 B(ApoB)mRNA 的 C-to-U 编辑使密码子 CAA(谷氨酰胺)变为 UAA(终止密码子),产生短链 ApoB48(肠道中),而未编辑的 mRNA 翻译为长链 ApoB100(肝脏中)。
总结
mRNA 的修饰是动态且多样化的,从两端的帽子和 poly (A) 尾,到内部的甲基化、假尿苷化及碱基编辑,共同构成了复杂的调控网络。这些修饰不仅影响 mRNA 的生命周期,还在发育、疾病(如癌症、神经退行性疾病)中发挥关键作用,是当前分子生物学和医学研究的热点领域。